


27 Nis SinirBilim 6.BÖLÜM – Duyu Sistemlerine Giriş
Bu bölümde duyu sisteminin ne olduğuna ufak bir giriş yapacağız. Ayrıca tat alma, koku ve somatosensori (dokunma, kas ve eklemlerin pozisyonu) sistemlerine de bu bölümde giriş mahiyetinde değineceğim onun dışında bu sistemler için ayrı konu açmayacağım.
Duyu sistemlerinin temel amacı Merkezi Sinir Sistemine (MSS) dış dünyanın bir temsilini vermektir. Duyu sistemlerinden aktarılan sinyaller algının oluşmasına ön ayak olurlar, depoladığımız anılarımızı harekete geçirerek bilinçli tecrübeye imkan tanırlar. Burada hissetmek ile algılamanın iki farklı şey olduğunu belirtmekte fayda var, şöyle bir benzetme yapabiliriz: duyusal sistemi aşağıdan yukarı çıkan bir mekanizma, algı mekanizmasını da yukarıdan aşağı inen bir mekanizma olarak düşünebiliriz. Aşağıdan yukarı çıkan mekanizma herhangi bir duyu organı bir şey algıladığında harekete geçer, gelen bu yoğun bilgi demeti arasından dikkatimizi hangisine vermemiz gerektiğini seçen mekanizma da yukarıdan aşağı inen mekanizma yani algı mekanizmasıdır.
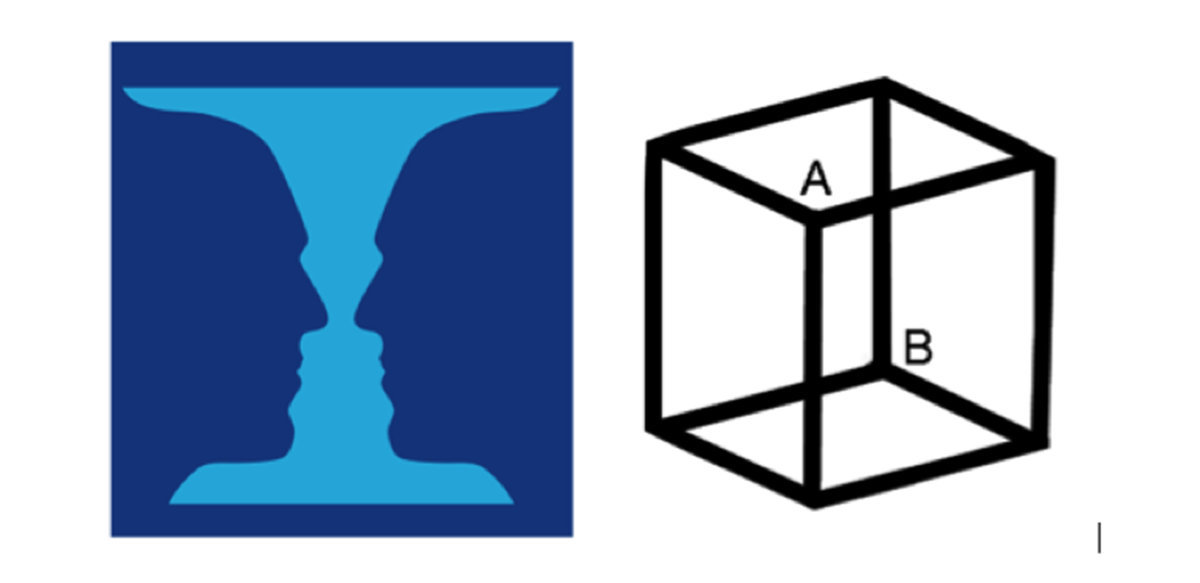
Bir örnek ile bu ikisi arasındaki farkın çok rahat anlaşılacağını düşünüyorum. Şimdi aşağıdaki resimdeki vazoya yada Necker Küpü olarak bilinen küpe bakalım. Vazoyu normal bir vazo olarak ve bir de insan figürü olarak görürüz. Şimdi bir de küpe bakalım. İlk bakışta B noktası küpün iç alt kısmındaymış gibi dururken biraz daha baktığımızda üst dış kısmında olduğunu da görüyoruz. İşte aşağıdan yukarı çıkan mekanizmadan gelen bilgi aynı olmasına rağmen, yani beyne gelen sinyal/veri değişmemesine rağmen algımız değişebiliyor.
Reseptörler
Çoğumuzun az çok bildiği gibi MSS’deki nöronlara veya omuriliğe dokunduğunuzda tepki vermezler, aynı şekilde ışığa yada sese de tepki üretmezler. Bütün bu fiziksel sinyallerin bir şekilde bütün nöronların anlayacağı dile/sinyallere dönüştürülmesi gerekir. İşte bu çevirme işlemini reseptör dediğimiz özelleşmiş hücre grupları gerçekleştirir. Her temel uyarı için (fiziksel, kimyasal, ısı, ışık vs.) özelleşmiş hücre grupları bulunur. Hatta her bir grubun içinde de sadece belli bir uyarı için daha da özelleşmiş hücreler bulunur, mesela sıcağı yada soğuğu algılayan hücreler gibi. Reseptörlerin bu farklı tepkilerinin sebebi tabii ki yapılarının ve kullandıkları kimyasalların farklı olmasından kaynaklanır.
Neredeyse bütün reseptörler algıladıkları enerjiyi membran voltajındaki değişime çevirirler (Bkz. Dinlenim ve Aksiyon Potansiyelleri). Yani algıladıkları enerjinin yoğunluğuna göre membran potansiyelini değiştirirler. Reseptörün görevi bu voltaj değişimini bir şekilde bağlı olduğu nöron topluluğuna ulaştırmaktır, bu hücre toplulukları genellikle gangliyon hücrelerdir ve bunların aksonları da ya beyne yada omuriliğe gider.
Reseptörlerin biçimleri ileride de göreceğimiz gibi duyu organlarına göre değişiklik gösterir, mesela işitsel sistemde saç hücreleri kullanılırken görsel sistemde çubuk ve konik reseptörler kullanılır.
Görsel sistemdeki reseptörler, ki aynı zamanda fotoreseptör olarak da bilinirler; retina, diensefalonun bir parçası olduğu için MSS’ne aittirler. Fakat işitsel, somatosensori ve koku sistemlerine ait reseptör ve gangliyon hücrelerin çoğu Çevresel Sinir Sistemi (ÇSS)ne aittir. Bu ufak gibi görünen fark büyük sonuçlar doğurur. ÇSS’ye ait olan reseptörler Schwann Hücreleri gibi bazı sinirsel olmayan yapılar tarafından desteklenirken fotoreseptörler gibi MSS’ye ait olan reseptörler sinirsel hücreler tarafından desteklenirler. Bu fark da fotoreseptörlerin ve bağlı oldukları gangliyon hücrelerin ileride de göreceğimiz gibi hasar gördüklerinde bir daha onarılamayacakları anlamına gelir.
Duyu sistemindekiler hariç diğer bütün reseptörler doğduğunuzda ne kadar ise o kadardır yani bunlara yeni reseptörler eklenmez. Hatta tam tersine yaşlandıkça reseptör sayısında ve bunun sonucu olarak duyusal keskinlikte azalmalar olur. Bu durum kendini 6 haftada bir yenileyen koku sistemi reseptörleri için bile böyledir, onlarda da yaş ilerledikçe yenilenme oranı ölüm oranını yakalayamaz.
Merkezi Yollar ve Bilgi İşleme
Gangliyon hücrelerden gelen aksonlar MSS’deki yolların başlangıcını oluştururlar. Görsel, somatosensori ve işitsel sistemdeki aksonlar talamusa ulaşmadan önce beynin orta kısmından karşıya geçerler, yani sağ gözden gelen aksonlar beynin sol yakasına geçerek sol talamusa ulaşır. Koku ve tat sistemlerinden gelen aksonlar ise bulundukları taraftan devam ederler.
Koku alma sistemi hariç diğer duyusal sistemlerden gelen aksonlar talamusu geçerek serebral kortekse doğru yol alırlar. Koku alma sistemindeki aksonlar talamusa uğramadan doğruca koku alma korteksine giderler.
Bu merkezi yollar genellikle ilerilerde bölünerek farklı işlevlere ayrılırlar bu olaya haritalanma denir. Mesela işitsel sistemdeki gangliyon hücreler MSS’ye girmeden alt parçalara bölünürler ve bu alt bölümler salyangoz nükleusun üç alt bölgesine ulaşarak son bulurlar. Her bir bölge farklı frekansa cevap verecek şekilde haritalanmıştır. Bu haritaların en çeşitlisi gözden gelen gangliyon hücreleri tarafından oluşturulmuştur. Bu gangliyon hücrelerden gelen aksonlar ileride de göreceğimiz gibi Lateral genikulat nükleusta altı ayrı katmanda haritalanırlar. Bu farklı haritaların görsel bilginin farklı özellikleri ile ilgilendikleri düşünülüyor. Ayrıca bütün bu farklı haritaların aynı anda paralel olarak işlenip bütünsel görsel/işitsel algının oluşturulduğuna inanılıyor. Bunları daha detaylı olarak görsel sistemi incelerken göreceğiz.
Aksonlar talamustan serebral kortekse gider demiştik. Aksonlar talamustan serebral kortekste gittikleri yerler genelde birincil korteksler olarak adlandırılırlar. Bu birincil bölgelerin doğrudan yada bir ara bölge ile bağlantı kurdukları bölgelere de bağlantı bölgeleri (association areas) denir.
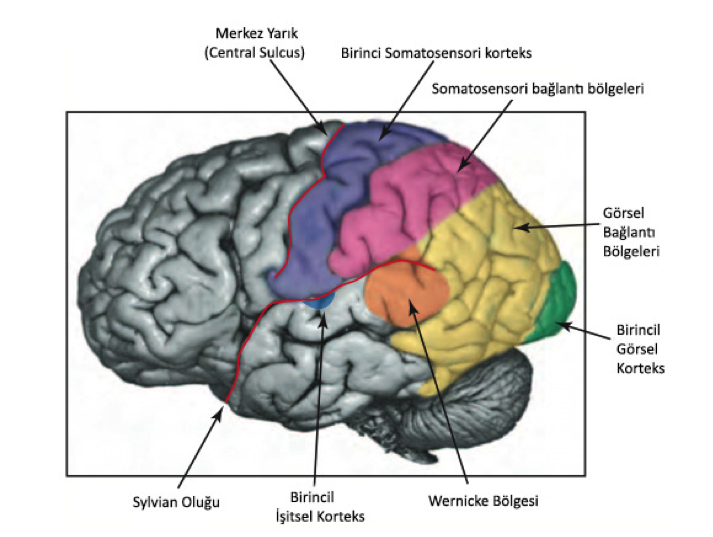
6.2 – Birincil Korteksler ve Bağlantı Bölgeleri
Duyusal korteksteki (ve serebral korteksin diğer çoğu bölgesinde) nöronlar altı katmanlı dizilmişlerdir. Orta katmanlar (3 ve 4) genel olarak talamustan gelen aksonların bitiş noktalarıdır. Birincil kortekslerde bu bölgeler daha büyüktür ve çok sayıda küçük nöron barındırır. Bu küçük nöronlar kum tanelerini andırdıklarından bu bölgeler tanecikli korteks bölgeleri (granular areas of cortex) olarak anılırlar.
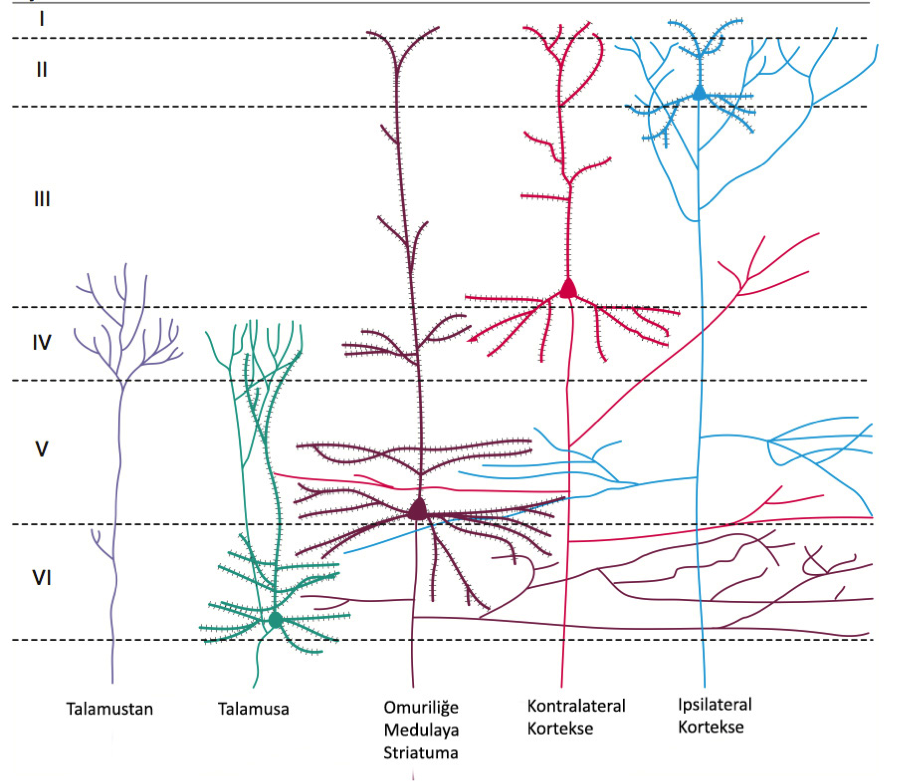
6.3 – Katmanlı Dizilim
Tat Alma
Her bir tat alıcısı (taste buds) yaklaşık 100-150 arası tat reseptör hücresi barındırır. Tat hücreleri epitelyumun (yani dilin dış zarının) sadece %1ini oluşturur. Tat alıcılarda ayrıca tat hücrelerini saran bazal hücreler bulunur, ayrıca tat alma aksonları bulunur. Normal bir insanda yaklaşık 2000-5000 arası tat alıcısı bulunur, bazı ekstra durumlarda bu sayı 500e inebilir diğer uçtaysa 20000e çıkabilir.
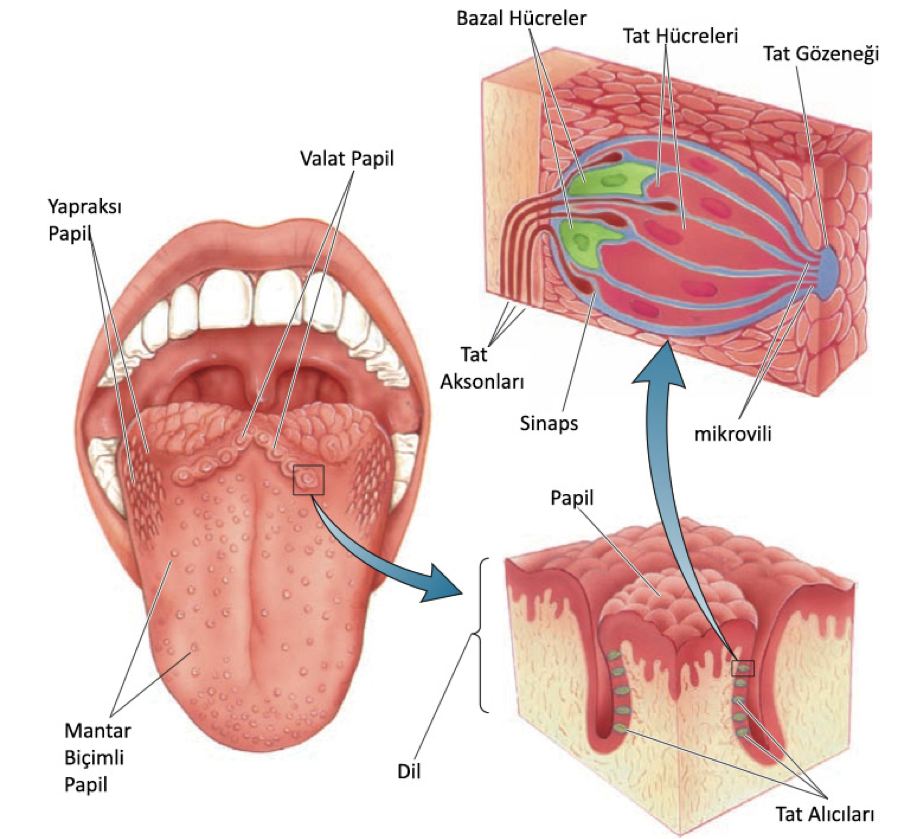
6.4 – Dilin Yapısı
Belli sınır yoğunlukların üzerinde her bir Papila sadece belli temel tatlara karşı duyarlı olurlar, yani dilimizde acıya duyarlı Papilalar ve tatlıya duyarlı Papilalar vs. vardır. Fakat yoğunluk arttığında çoğu Papila’nın seçiciliği düşer yani aşırı yoğun ekşiye maruz kalan tatlıya-duyarlı Papila bu ekşiye tepki verir. Bu seçicilikteki göreceli eksiklik/düşüş duyusal sistemlerin ortak özelliğidir. Çoğu duyusal reseptör uyarıcı ayrımı yapmazlar. Bu durum bir paradoks oluşturur: eğer tek bir tat alıcısı dondurma ile muz uyarılarına verdiği tepkiler çok az değişiyorsa nasıl oluyor da iki farklı çikolata arasındaki farklı anlayacak kadar keskin tat alabiliyoruz? Cevap tabii ki beyinde gizli.
Burada gizli derken gerçek manasını kastettim yani harbiden gizli henüz nasıl olduğu net bilinmiyor yani.
Tat Alıcı Hücreler
Tat alıcı hücrelerin kimyasal tepki veren kısmı dilin yüzey kısmında bulunan ufak bir kısımdır ve tepe ucu (apical end) olarak adlandırılır. Bu tepe uçlarda mikrovili adı verilen ince uzantıları bulunur (Resim 6.4). Tat alıcı hücreler nöron değildirler fakat bu tat aksonları ile bağlantı/sinaps kurarlar. Tat alıcıları normal bir büyüme-ölüm-yenilenme döngüsüne sahiptirler, bir tat hücresinin ömrü yaklaşık 2 haftadır. Bu süreç bağlı bulunan sinir hücresi tarafından değiştirilebilir, eğer bağlantı kesilirse tat alıcısı yok olur.
Tat alıcı hücresi uygun bir kimyasal tarafından harekete geçirildiğinde dinlenim potansiyeli (genelde depolarize olarak) değişir. Bu voltaj değişimine reseptör potansiyeli denir, eğer bu potansiyel yeterince kuvvetliyse çoğu tat alıcı hücre nöronlar gibi ateşleme yapabilir. Reseptör potansiyelinin depolarizasyonu voltaja-duyarlı kalsiyum kanallarının açılmasına ve Ca2+’ın içine girmesine ve iletici moleküllerin salınımının tetiklenmesine sebep olur (2 ve 3. bölümde gördüğümüz gibi). Bu tat hücresinden aksona yapılan temel bir sinaptik iletimdir. Tat reseptörlerinin transmiterleri henüz bilinmiyor ama bu transmiterlerin postsinaptik nöronda aksiyon potansiyeline sebep olarak tat sinyalini beyin köküne ilettiğini biliyoruz.
Tat Dönüşümünün Mekanizması
Çevreden gelen bir uyarının duyusal alıcı hücrede elektriksel sinyale dönüşmesine transdüksiyon/dönüşüm (Elektronik ile uğraşanlar transduser olarak da bilirler) denir. Sinir sisteminde çok sayıda dönüştürme mekanizması bulunur; böylece sese, kimyasallara, ışığa, basınca duyarlı olabiliriz. Bu dönüşümün mekanizması duyusal sistemin de neye duyarlı olacağını belirler, yani duyusal sistem ışığa duyarlıysa dönüştürme mekanizması da ona göredir. Görürüz çünkü gözlerimizde fotoreseptörler bulunur, eğer fotoreseptörler dilimizde olsaydı belki dilimizle görebilirdik.
Bazı duyusal sistemler tek bir reseptör türüne ve tek bir dönüşüm mekanizmasına sahiptirler. Fakat tat dönüşümünde birkaç mekanizma birden kullanabilir:
- (a) doğrudan iyon kanallarından geçerek (tuzlu ve ekşi),
- (b) iyon kanallarını engelleyerek (ekşi),
- (c) G protein reseptörlerinin iyon kanallarını harekete geçirmeleri sayesinde (acı, tatlı, umami).
Bunlar daha önce gördüğümüz normal sinyal mekanizmalarıdır.
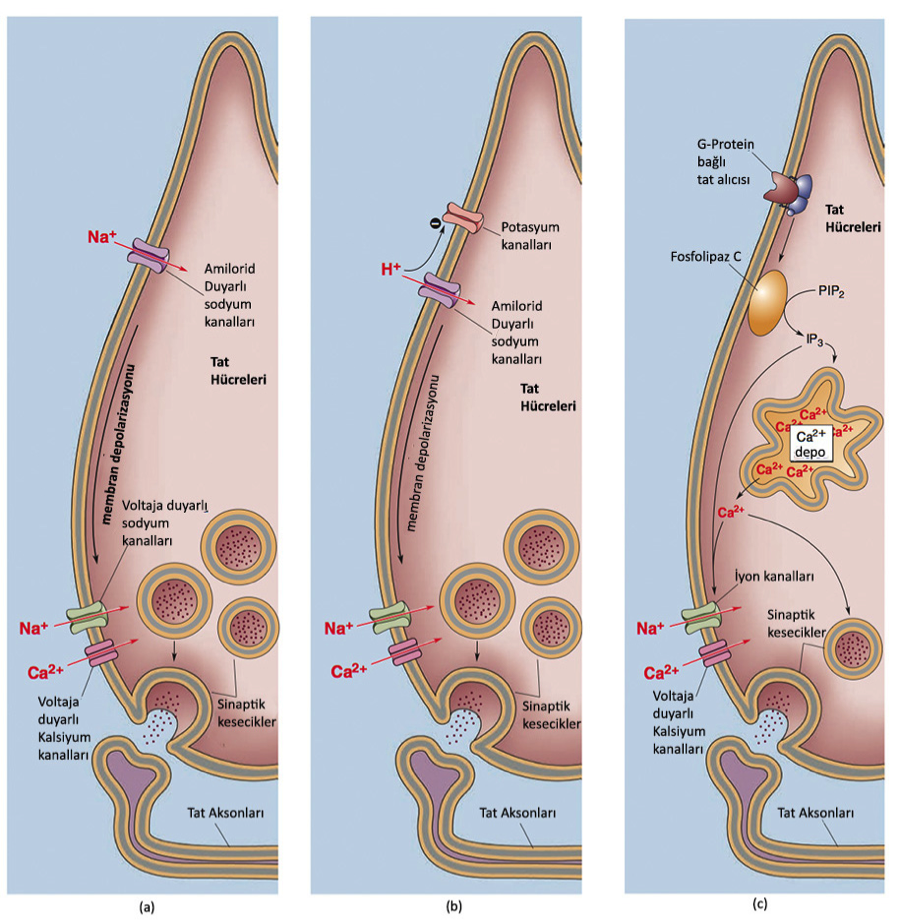
6.5 – Tat Alma Mekanizmaları
Tuzluluk
En bilinen tuz sudan sonra kanın, denizin ve çorbanın temel bileşeni olan sofra tuzudur (NaCI). Tuzun tadı aslında genelde katyonun (Na+) tadıdır ve algılanması için yoğunluğunun fazla olması gerekir. Tuza duyarlı tat hücreleri özel Na+ duyarlı kanallara sahiptirler. Tat kanalları voltaja duyarsızdır ve sürekli açıktırlar. Eğer tuzlu çorbayı içerseniz, hücre dışında Na+ yoğunluğu artar, bundan dolayı Na+ içeri akmaya başlar böylece hücre depolarize olur. Bu depolarizasyon voltaja duyarlı sodyum ve kalsiyum kanallarının açılarak nörotransmiterlerin tat aksonlara salınımına sebep olur.
Ekşilik
Yiyeceklerdeki ekşiliğin sebebi içerdikleri yoğun asittir (düşük pH). HCl gibi asitler suda çözülür ve hidrojen iyonları üretirler (proton yada H+) yani Protonlar asidin ve ekşiliğin temel kaynağıdır. Tat reseptörlerini iki şekilde etkileyebilirler: ilk olarak H+ amilorid-duyarlı sodyum kanallarından geçer ve hücreyi depolarize eder. İkinci olarak da hidrojen iyonları K+ duyarlı kanalları bloklayabilir. Kanalın K+ geçirgenliği azaldığında hücre depolarize olur. Bunlar muhtemelen tek ekşi dönüşüm mekanizmaları değil çünkü bunların dışında pH değerindeki değişim de bütün hücresel süreçleri değiştirebilir.
Acı
Acı dönüştürme mekanizmaları hakkındaki bilgilerimiz 2000 yıllarında bulunan iki gen ailesi (T1R ve T2R) sayesinde epey arttı. Nesnenin acılığı 30a yakın farklı T2R alıcısı tarafından algılanır. Acı alıcıları zehir algılayıcılarıdır aslında. Son yapılan araştırmaların gösterdiğine göre acı, tatlı ve umami alıcılarının hepsi aynı 2. taşıyıcıları (2. messenger) yollarını kullanıyorlar. Eğer tastant acı alıcısına tutunursa, G-proteinini harekete geçirir, buda Fosfolipaz C enzimini tetikler buda inositol trifosfat (inositol triphosphate IP3) hücre içi taşıyıcısının üretimini artırır. Tat hücrelerinde IP3 özel bir iyon kanalını harekete geçirip açılmasını ve Na+’ın içeri dolasına sebep olur ve hücre depolarize olur. Depolarizasyon da voltaja duyarlı kalsiyum kanallarının açılmasını ve Ca2+’ların hücre içine girmesine neden olur. Na+ ayrıca hücre içindeki belirli bölgelere de Ca2+ salınımını tetikleyebilir, bu iki Ca2+ kaynağı nörotransmiter salınımını tetikler.
Tatlılık
Tat dönüştürme işlemi de acıya benzerdir. Bundan farklı olarak bu süreçte tatlı alıcıları bulunur ve bu alıcılar T1R ailesinin iki önemli üyesine ihtiyaç duyarlar: T1R2 ve T1R3, eğer bunlardan biri yoksa yada değişime uğramışsa canlı tatlı şeyleri algılayamayabilir.
Merkez Tat Alma Yolları
Yukarıda da gördüğümüz gibi tat bilgisi tat alıcılarından birincil tat aksonlara oradan da beyin köküne ve talamusa ve serebral kortekse kadar gider.
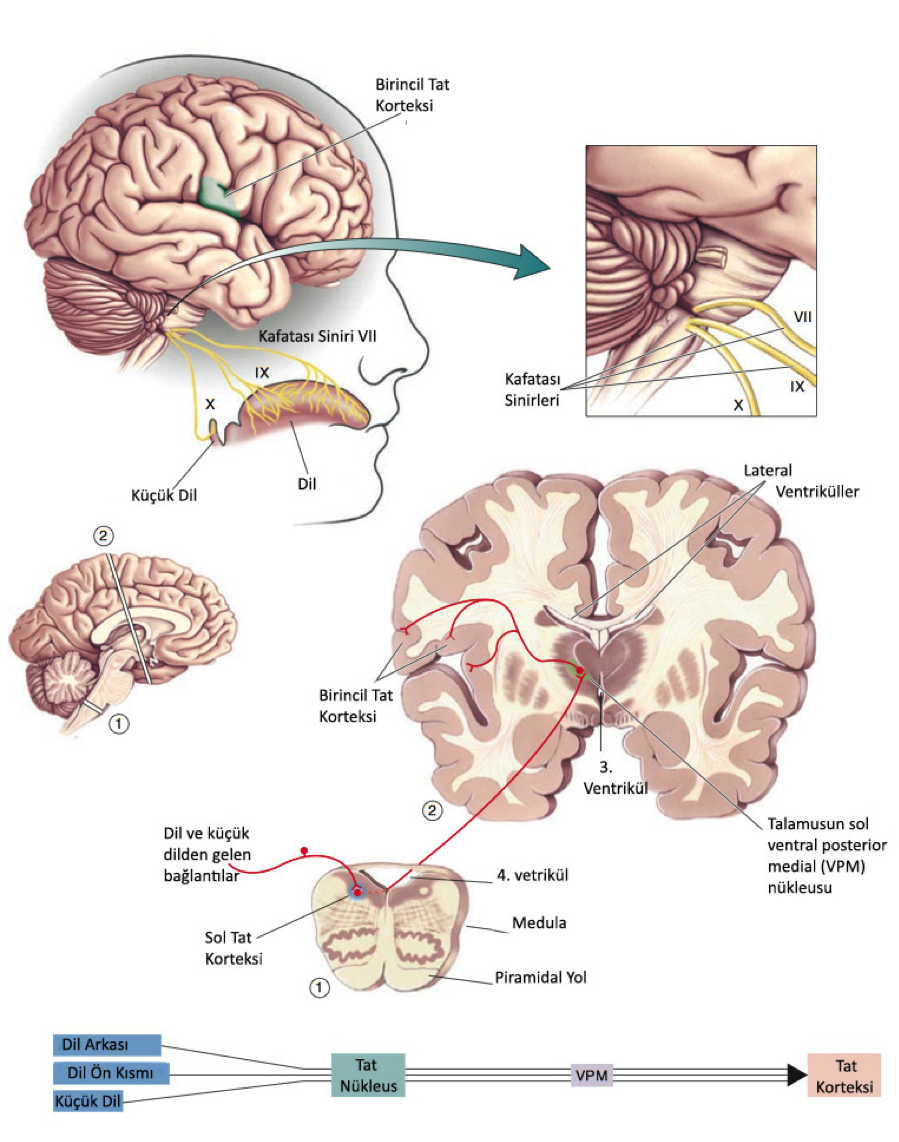
Resimden de görebildiğimiz gibi tat bilgileri dilden kafatası sinirleri tarafından medulaya taşınır. Tat aksonları meduladaki tat nükleusa gider. Tat nükleusundaki aksonlar talamustaki nöronlara bağlanırlar ve oradan da serebral korteksteki ilgili bölgelere giderler.
KOKU
Kokunun ne işe yaradığını hepimiz az çok biliyoruz o yüzden direk konuya dalıyorum.
Koklama Organı
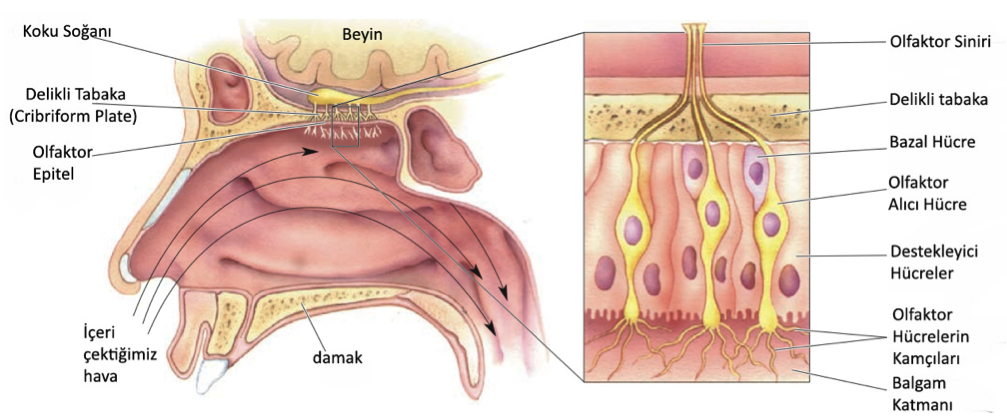
6.7 – Koklama Organı
Aslında burnumuzla değil de burnumuzdan çok daha ufak ince bir hücre tabakası olan olfaktor epitel ile koku alırız. OE’de üç temel hücre çeşidi bulunur. Olfaktor alıcı hücreler dönüşümün merkezidirler. Tat hücrelerinin aksine olfaktor hücreler gerçek nöronlardır, ve MSS’ye giden kendi aksonları bulunur. Destekleyici hücreler gliyaya benzerler diğer görevlerin yanında balgam üretimine de yardım ederler. Bazal hücreler yeni alıcı hücrelerin kaynağıdırlar. Olfaktor hücreler (tat hücreleri gibi) büyür ölür ve yenilenirler, bu döngü 4-8 hafta sürer. Aslında bu hücreler MSS’de bulunan ve yenilenebilen nadir nöronlardandır.
Havayı içinize çektiğinizde, havanın sadece az bir kısmı olfaktor epitel ile temas kurar. Epitel ince katmanlı bir balgam yayar. Odorant diye adlandırılan kimyasal alıcı hücrelere ulaşmadan uyarı, bu balgamda çözülür. Bu balgamda su, şeker, antikor enzimler gibi çeşitli proteinler ve tuz bulunur. Antikorlar özellikle önemlidir çünkü bazı virüsler direk olfaktor hücreler vasıtasıyla beyne ulaşabilirler.
Olfaktor Alıcı Hücreler
Olfaktor alıcı nöronlar epitele kadar uzanan tek ve uzun bir dentrite sahiptir. Epiteldeki ucundan yayılan birkaç uzun kamçı bulunur. Balgamda çözülen odorantlar bu kamçıların yüzeyine tutunurlar ve dönüşüm sürecini başlatırlar. Alıcı hücrenin diğer tarafında ise ince miyelinsiz akson bulunur.
Olfaktor aksonlar koku soğancığına giderler. Olfaktor aksonlar çok kırılgandır ve hasar gördüklerinde anosmi adı verilen koku alma yitimi hastalığı ortaya çıkar.
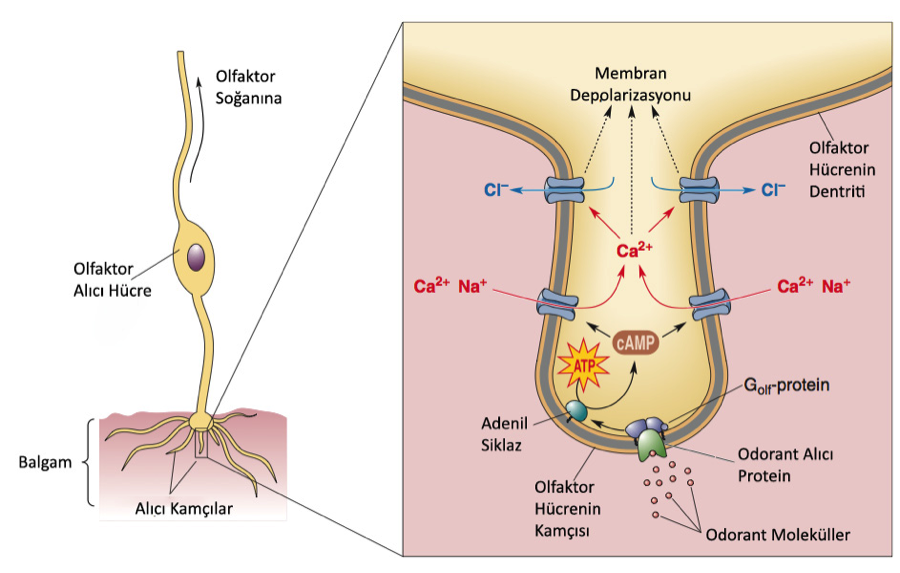
6.8 – Olfaktor Alıcılar ve Dönüşüm
Olfaktor dönüşümün izlediği yol şu şekilde özetlenebilir:
Odorant moleküller odorant alıcı proteinlere tutunurlar. G-protein (Golf) uyarılır. Adenil Siklaz harekete geçer. cAMP oluşturulur. cAMP belirli katyon kanallarına tutunur. Kanallar açılır ve Na+ ve Ca2+ içeri dolar. Ca2+ ile aktif olan klorid kanalları açılır. Membran potansiyeli gerçekleşir. İşte hiç beklemediğiniz bir anda saçma sapan bir sabun molekülü epitelden girip bu bir ton süreçten geçip sizi, yan komşular halı yıkarken sizin maç yaptığınız sokağa, çocukluğunuza, götürebilir, sonra duygulanırsınız, dalarsınız hey gidi günler dersiniz ama tabii dersi kaynatmayalım şimdi bunlar başka yazıların konuları.
Merkezi Olfaktor Yolları
Olfaktor alıcı nöronlardan iki koku soğanına akson gider. Bu soğancıklar sinirbilimcinin harikalar diyarıdır, çünkü muhteşem dentritik dizilimlere sahip sinirsel devrelerle, ve çok çeşitli yüksek derece nörotransmiterlerle doludurlar. Her bir soğanın girişinde yaklaşık 2000 tane glomerül (glomeruli) adında yuvarlak yapılar bulunur. Her bir glomerülde reseptör hücrelerden gelen yaklaşık 25000 olfaktor akson ucu bulunur.
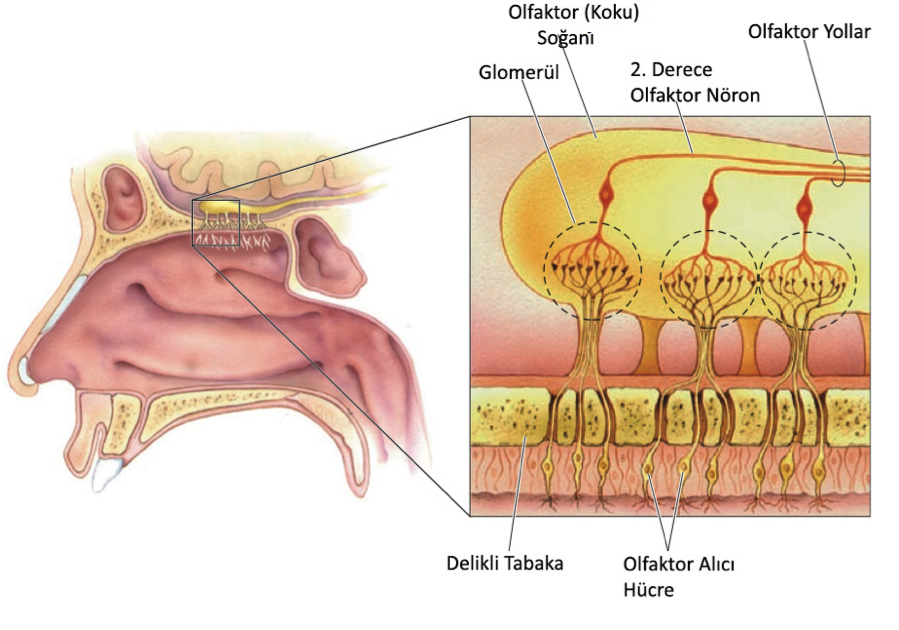
6.9 – Koku Yolları
Şekilden de görebildiğimiz gibi olfaktor alıcı hücrelerin aksonları delikli tabakadan geçerek olfaktor soğanına girerler. Birkaç dala ayrıldıktan sonra her bir olfaktor aksonu glomerülün içinde 2.derece nöronlarla bağlantı kurar. Bu 2. Derece nöronlar da olfaktor yollar aracılığıyla beyne aksonlarını yollarlar.
Çoğu beyin bölgesinde olfaktor bağlantı vardır. Koku soğanından çıkan aksonlar olfaktor yollardan geçerek doğrudan birkaç noktaya bağlanırlar. Bu noktalardan en ilginci serebral korteksin ilkel bölgelerinden biri olan olfaktor kortekstir.
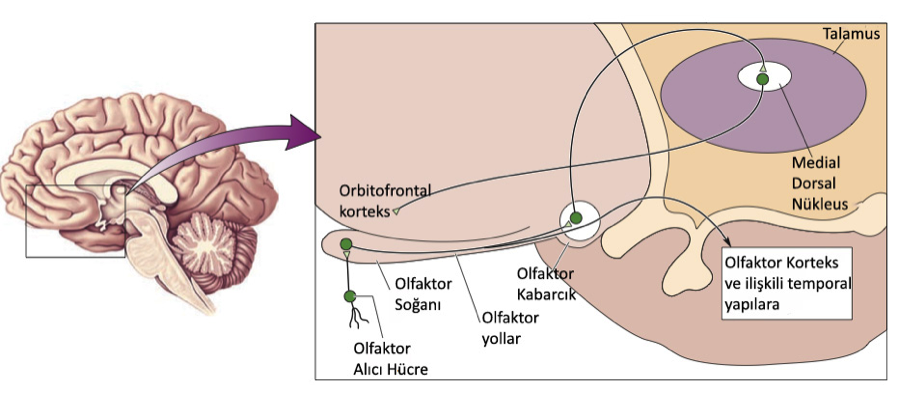
6.10 – Olfaktor Yollar
Yukarıda koku aksonlarının izledikleri yolları ve uğradıkları beyin bölgelerini görebiliriz. Bilinçli koku alma muhtemelen bu bölgelerde ortaya çıkıyor.
Koku sisteminde de tat alma sistemine benzer bir paradoks bulunur. Her bir reseptör kendine has uyarılara tepki verir, yani her bir hücre çok çeşitli kimyasallara tepki verebilir. Fakat biz bu kimyasalları kokladığımızda bunları ayırt edebiliriz. Peki tek bir hücrenin yapamadığını beyin nasıl yapıyor? Burada üç önemli görüşü özetlemek istiyorum: (1) her bir koku geniş nöron topluluğu tarafından temsil edilir; (2) belli kokulara tepki veren nöronlar uzamsal haritalarda temsil ediliyor olabilir; (3) aksiyon potansiyellerinin zamanlamaları belli kokuların kodlanmasında kullanılıyor olabilir.
Burada bu görüşlerin detaylarına yer vermeyeceğim ve bu konuyu burada noktalayacağım.
Hiç yorum yok.